
Tabla de contenidos
Del punto omega de Teilhard a la neoortogénesis de la nueva biología (y Parte II)
Por Juan Luis Doménech Quesada

Principio de mutación
Tal y como deja claro Lynn Margulis en todas sus obras, la simbiosis y la cooperación (fusión o unión) tiene mucha más importancia en la evolución que la mutación al azar y la acción disgregadora de la selección natural. Margulis y Sagan (1999) proponen una primera fusión entre las arqueobacterias fermentadoras y las eubacterias nadadoras para dar lugar a los protocistas anaeróbicos; una segunda fusión entre estos y las eubacterias aeróbicas para dar lugar a las células con núcleo; y una tercera fusión entre estas y las cianobacterias fotosintéticas para dar lugar a las algas unicelulares. En la actualidad, se ha impuesto la idea de que todos los animales superiores, entre los que se encuentra el hombre, proceden de una gran mutación por endosimbiosis.
Si bien la evolución gradual por selección natural tiende a la disgregación de las partes, la Evolución progresiva por saltos consiste precisamente en lo contrario: la unión de las partes. Parece evidente que la unión de las partes (dos células sin núcleo) dio lugar la célula con núcleo. Parece evidente que la unión de las partes (dos células, o más, con núcleo) dio lugar a los pluricelulares. Y empieza a ser evidente que la unión de las partes dio lugar, incluso, a los vertebrados: en este caso, por medio de genes duplicados en el genoma y posteriores mutaciones independientes. La evolución de los vertebrados por poliploidía está prácticamente demostrada merced a los últimos avances de la biología molecular. Parece ser que los invertebrados sólo poseen una copia de genes Hox, mientras que los peces sin mandíbula poseen una doble copia producida por duplicación génica.
Los peces mandíbulados poseen cuatro complejos Hox, producida por duplicación de la anterior, al igual que los tetrápodos derivados de estos, todos los cuales poseen cuatro copias. Se piensa que estas poliploidías continuaron y que los ancestros de los peces teleosteos tenían 8 complejos Hox, lo que explicaría por qué algunos peces actuales poseen esas 8 copias (como el pez cebra). La mayor parte de los peces actuales poseen cuatro copias, obtenidas por pérdida de la mitad de las ocho copias ancestrales. Una vez más, la reducción de genes, más frecuente de lo que se cree, va acompañada de simplificación y regresión morfológica, tal y como sucede con el pez Fugu, en el que se reduce la pelvis y las costillas (Aparicio, 2000).
La teoría de la mutación que antaño defendieran Hugo de Vries, o la importancia que le diera William Bateson (con su teoría de la “mutación negativa” o inhibición de genes), J.P. Lotsy (y su teoría de la “hibridación creativa”) o Goldschmidt (autor de la teoría del “monstruo prometedor”), entre muchos otros, fue recuperada recientemente por Stephen Jay Gould, el brillante paleontólogo de Harvard que redescubrió, junto con Niles Eldredge, que los antiguos científicos tenían razón (Gould, 1994). Su teoría del equilibrio puntuado, según la cual la evolución consiste en cambios bruscos seguidos de grandes períodos de estabilidad, redescubre el saltacionismo de antaño, cuyo nombre reivindicamos, en honor de tan ilustres científicos.
Tanto la teoría de la “integración de sistemas complejos”, de Máximo Sandín, como la teoría endosimbionte y la simbiogénesis, de Margulis y Sagan, dan una vuelta de tuerca al proceso iniciado por Gould: ¿qué está ocurriendo durante esos saltos evolutivos?; ¿qué fenómenos podrían dar lugar a esa “evolución rápida” claramente observable por los paleontólogos?.
Dejando a un lado los grandes cambios provocados por mutaciones ocurridas en las primeras fases de la ontogenia en formas dotadas de plasticidad fenotípica (finalmente reconocidas por los modernos biólogos del desarrollo), la base de la teoría de Sandín (lamarckista, por cierto) y su equipo de la Universidad Autónoma de Madrid, parte de la capacidad de los virus para insertarse en el genoma, en una especie de “módulos compactos”, con funciones perfectamente definidas y operativas. Sus observaciones le llevan a concluir que “los genomas animales y vegetales están constituidos por una suma de genomas bacterianos y virales” (Sandín, 2003). Si observamos algunos de los árboles evolutivos más recientes (figura 9), podemos darnos cuenta de cómo nos estamos alejando de los dogmas que hasta ahora han imperado en las ciencias de la vida.
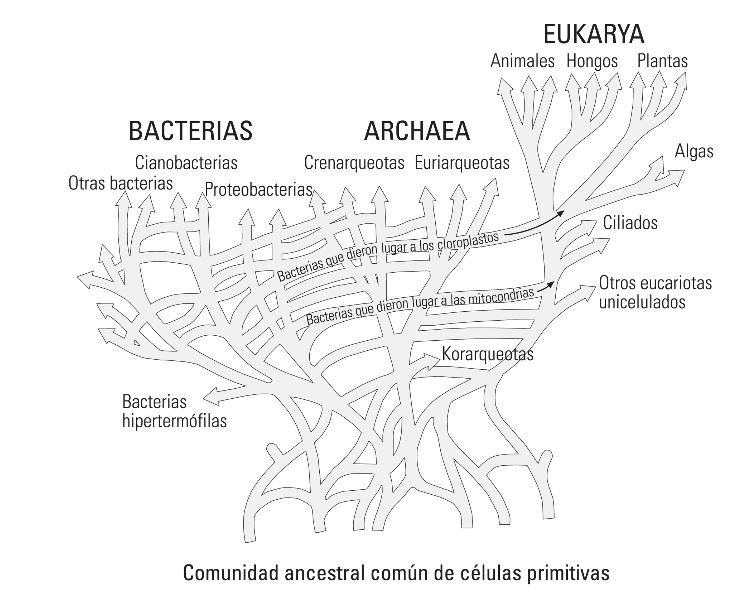
Figura 9. Versión «en red» del árbol de la vida, de W. Ford Doolittle (1999)
La Versión «en red» del árbol de la vida, de W. Ford Doolittle (1999), donde se muestra que los eucariotas adquirieron los cloroplastos y mitocondrias de las bacterias, así como la universal transferencia horizontal de ADN. Aunque rompe moldes, sigue integrando los árboles en pirámide de Gould con los clásicos en abanico.
Es decir, la evolución por saltos podría estar basada, por ejemplo, en “infecciones” masivas de módulos de ADN (vía virus) acaecidas ante especiales eventos ambientales movilizadores de energía (grandes cambios geoclimáticos). Cada uno de estos módulos contendría uno o más algoritmos genéticos perfectamente funcionales, que permitiría acotar bastante más los cálculos cronológicos que nos ofrece una evolución meramente azarosa y micromutacional, al tiempo que permitiría obviar, de una vez por todas, la persistente carencia de formas intermedias en el registro fósil. Como ya hemos dicho, un muy sugerente mecanismo de evolución cuántica es la duplicación genética y posterior integración de genes en una de las copias (libre para «experimentos» naturales de todo tipo), mientras que la otra copia mantendría intactas las funciones originales.
Son ya decenas las citas y sugerencias acumuladas, de las que entresacamos una al azar del mencionado artículo de Sandín (2003):
“Existen sintagmas (complejo genproteina) que controlan el desarrollo embrionario de, por ejemplo, ojos, patas, alas…, independientemente del tipo de ojo, pata o ala, es decir del Phylum al que correspondan: los apéndices de vertebrados y artrópodos no son estrictamente órganos homólogos pero vemos que en su morfogénesis hacen uso de genes y sintagmas conservados” …… “Los genes Hox, implicados en el control del desarrollo embrionario de tejidos y órganos, son, como todos sabemos, secuencias repetidas en tándem, y sabemos también que los responsables de las repeticiones en el ADN son los retrotransposones” (a propósito de una publicación de García Bellido sobre la gran “explosión cámbrica”).
Y otra cita de uno de los últimos libros de Margulis y Sagan (2003), cuyo título lo dice todo (Captando genomas):
«Las fusiones biológicas que se inician en forma de simbiosis, constituyen el motor de la evolución de las especies…. demostraremos aquí que la fuente principal de variación hereditaria no es la mutación aleatoria, sino la adquisición de genomas«. Recomendamos la lectura completa de este libro para comprobar si lo demuestra o no, y cerrar así, de forma definitiva, este Principio de Mutación.
La activación-desactivación de genes y de complejos de genes “algorítmicos” queda patente con el reciente descubrimiento de que «el insecto palo” ha perdido y recuperado sus alas varias veces en el curso de la evolución. En esta última ocasión, las alas se recuperaron a través de instrucciones genéticas que permanecieron latentes durante….. ¡50 millones de años! (Whiting et al., 2003).
Entre los insectos existen numerosos casos de pérdida de alas (con tendencia al parasitismo) en heterópteros, homópteros, anopluros, malófagos, psocópteros, estrepsípteros, himenópteros, sifonápteros, etc. Entre las cochinillas o cóccidos «las especies más evolucionadas ofrecen una serie de regresiones morfológicas, como pérdida de patas, alas, antenas, ojos, a menudo piezas bucales, todos los machos de cóccidos están desprovistos de estiletes y rostro; el orificio bucal no es funcional» (Grassé, 1976). Ha sido demostrada la relación de simples genes con importantes caracteres morfológicos y grupos de caracteres, como el gen Hand2 (pez cebra, ratones, pollos…) que codifica para una proteína que regula la expresión de genes sin los cuales no se desarrolla la placa lateral mesodérmica, ni las estructuras que derivan de él: mandíbulas, corazón o aletas pectorales.
Las macromutaciones, el saltacionismo y los «monstruos prometedores» de Goldsmitch han sido demostrados, y con estos, lo fácil que resulta incrementar la complejidad de forma súbita o regresar a estados pasados. Algunos críticos creen que regresión significa volver por los mismos pasos evolutivos ya recorridos. Diremos, para su satisfacción, que la regresión es todo lo contrario: es avance, pero avance por el camino contrario al de la complejidad, es decir, el de la simplificación.
Principio de dirección
Es mucho lo que hay que decir, desde los experimentos de Cairns, sobre la direccionalidad evolutiva para resumirlo en un corto artículo. El equipo de Cairns observó que en situaciones de estrés ambiental, la levadura presentaba la mutación necesaria para su adaptación, en mayor frecuencia de la esperada.
Como uno de los posibles mecanismos para explicar este comportamiento, aparentemente lamarckiano, sugería la producción, a partir de un único gen, de varios mRNA, para luego fijar en el DNA el mejor resultado adaptativo, por transcripción inversa (Cairns et al., 1988). Aunque algunos lo llaman simplemente “mutación adaptativa”, José Leonel Torres, en una obra más reciente (1995), revisa toda esta discusión desde los experimentos de Luria y Delbrück de 1943 (con los que se reabre éste nuevo viejo debate) y la interpretación de J. Mittler y R. Lenski (1990), en respuesta a la de Cairns, Overbaugh y Miller (1988), concluyendo:
«Una posibilidad considerada por Cairns y colaboradores, es la producción de múltiples copias no idénticas de mARN por un mismo gen, bajo condiciones de estrés. Cada una de ellas originaría una enzima distinta, y si la célula poseyera la capacidad de medir la eficacia de sus productos y de copiar en inversa sobre el genoma, por la reversotranscriptasa, solo aquella versión de mARN que produjera la mejor proteína, aceleraría precisamente las mutaciones útiles en la solución de su problema fisiológico… Un mecanismo como el descrito conferiría una gran ventaja adaptativa a los organismos que lo poseyeran, frente a aquellos que sólo mutaran aleatoriamente. Contradiría, según dijimos, una de las hipótesis básicas en la teoría de evolución de Darwin, reintroduciendo el lamarckismo en la biología; permitiría el flujo de información del ambiente al genoma«.
Pues bien, solo diremos que este mecanismo ya se ha encontrado (Herbert et al., 1999), sin que aparentemente la comunidad científica haya reaccionado en absoluto. Todo lo contrario, y esto es lo más alarmante, algunos especialistas ignoran aspectos, como que «los rasgos adquiridos pueden serlo no como tales, sino como genomas» (Margulis et al., 2003), e insisten en que «no existe ningún fenómeno biológico concebible capaz de apoyar la evolución lamarckiana» (Landman, 1993).
Un reciente y curioso artículo sobre la evolución de los lagartos del Caribe, parecen indicar que la evolución no tiene tantos caminos para elegir, sino que las direcciones están más marcadas de lo que parece. Resulta que estudiando la evolución de los lagartos en varias islas diferentes del Caribe, los autores llegaron a la conclusión de que a partir de un fundador generalista, se forman diferentes formas especializadas de lagartos, unos adaptados a vivir en las ramas, otros en el tronco, otros en la cúpula arbórea, etc., es decir, lo que siempre sucede, como acabamos de ver con los pinzones de Darwin. Pero, lo original de este caso es que ocurre lo mismo en 4 islas diferentes y afecta a comunidades enteras y no sólo a una especie como en los clásicos ejemplos de convergencia evolutiva (Losos et al., 1998).
Este ejemplo ha reabierto entre los zoólogos el viejo debate acerca de si existe un cierto determinismo evolutivo, es decir, ¿discurre la evolución al azar o existen en realidad unos pocos caminos donde elegir? Pero, lo cierto es que el debate nunca se había cerrado entre los biólogos del desarrollo, los cuales descubrieron ya hace tiempo que existen «canalizaciones» que restringen el camino a seguir, es decir, en cierto modo descubrieron el famoso «programa interno» de los antiguos ortogenistas que muchos expertos despistados aún se niegan a admitir.
Las llamadas «tendencias progresivas» de Haeckel tienen su continuidad en autores modernos como Brian Goodwin (1998), que sugiere tendencias en el incremento de complejidad debidas a la morfogénesis autoorganizada de los sistemas; o Michael McKinney (1998), que no solo defiende las ideas de progreso e incremento de complejidad, sino que propone un «progreso evolutivo esencial absoluto«, basado precisamente en ese incremento histórico de complejidad. Es también el «progreso global» de Daniel Dennett, o la «gran dirección» de David Hull. Como dice McKinney:
«la evidencia paleontológica indica que los patrones de desarrollo de todas las formas de vida se han ido haciendo más restrictivos….. se ha reunido una considerable evidencia ontogenética y paleontológica a favor de que el diseño corporal de los diversos grupos de organismos se han ido congelando gradualmente tras un período inicial de relativa plasticidad».
Podemos especular lo indecible con este tema, pero no es necesario. Basta con analizar los hechos ya acontecidos. Si volvemos a nuestro árbol de la vida, aunque ahora representado en forma de flecha (figura 10), vemos que el fenómeno evolutivo más espectacular y, a la vez, el más ignorado por la inmensa mayoría de los evolucionistas, es lo que hemos llamado «exclusividad del progreso absoluto»: ¿por qué no hay más de una rama que presente varios saltos de complejidad? Es más, ¿por qué no existen numerosas ramas que presenten dicho comportamiento, como cabría esperar de una evolución al azar? ¿Por qué no se han formado eucariotas u otros tipos de células complejas, múltiples veces? ¿Por qué no se han formado vertebrados u otra cosa partir de varios tipos de invertebrados?. Hace poco más de dos décadas, aun se pensaba que la mayoría de los grupos eran polifiléticos y que los organismos pluricelulares se habrían desarrollado independientemente en unas 17 ocasiones, por lo menos, a partir de antecesores unicelulares (Valentine, 1979).
Era lo lógico. Pero, hoy en día, la cosa se ha reducido a tres: las plantas, los animales y los hongos, e incluso a dos (animales y hongos por un lado, y plantas, por otro), tal y como piensa L. Margulis. Incluso, en virtud del principio de exclusión, no hay que descartar un único origen de los seres multicelulares, separándose posteriormente los animales y plantas (bien por adición de cloroplastos, o bien por pérdida de los mismos en una de las líneas filogenéticas).

Figura 10: La «flecha de la vida»
La «flecha de la vida» muestra con claridad los cuatro principios (C: niveles de complejidad): a) de progreso (C1 a C11), b) de involución o selección (C1 a C1; C2 a C2, etc.), c) de salto (C1 a C2; C2 a C3, etc.) y d) de dirección (exclusividad de la línea C1 a C11).
Las bacterias sólo han dado lugar a un nuevo nivel de complejidad en una única ocasión y desde hace 3.500 millones de años, no han creado nada nuevo. ¿Por qué?. Quizás por casualidad, pero es que exactamente lo mismo ha ocurrido con los siguientes niveles de complejidad. Demasiada casualidad.
Concluyendo: las posibilidades evolutivas no son tan elevadas como se pensaba; la historia de la vida deja entrever un importante componente determinista; y la «gran dirección» es una clara ortogénesis retrospectiva y excluyente que solo puede tener dos explicaciones naturales: una muy improbable casualidad, o la existencia de algún tipo de autoorganización que, además de incrementar la complejidad de forma constante, introduzca algún tipo de exclusión que impida nuevos saltos de complejidad en el resto de las ramas evolutivas.
No podemos extendernos aquí en nuestra opinión sobre las causas de tal comportamiento evolutivo (la «reina Alicia»; Doménech, en preparación), ni en las de autores como Agudelo y Alcalá, basadas en las teorías de sistemas complejos (Sandín et al., 2003), pero son en ellas en las que basamos nuestra esperanza de que las antiguas teorías finalistas se puedan sustituir por una nueva síntesis mucho más amplia que la anterior, y de corte ortogénico, para la que proponemos el nombre de neoortogénesis.
Aunque la mayor parte de los evolucionistas han optado por esconder la cabeza debajo del ala, el primer paso para encontrar la explicación adecuada es ver con claridad el árbol de la vida. En este artículo nos hemos ceñido a los hechos revelados por el registro fósil y desvelados, poco a poco, por los científicos más sagaces. Quizás el que más ha destacado, y hay que decirlo como pertinente y merecido homenaje, ha sido Teilhard de Chardin, con su teoría, ortogénica y finalista, del punto omega, el cual, aunque con interpretaciones más discutibles, vio con lucidez unos hechos que para muchos se nos antojan inapelables.

Figura 11. Si queremos, podemos sustituir nuestros viejos prejuicios «antropocéntricos» por nuevas concepciones, «bacteriocéntricas» por ejemplo , donde nos parezca que el Homo sapiens no es más que una bacteria «un poco evolucionada». Sin embargo, el árbol aquí representado es exactamente el mismo que el de la figura anterior («la flecha de la vida») y que el de la figura 4.
Necesitamos nuevas mentes para nuevas visiones (C1, C2, C3, etc. : niveles de complejidad).
El reto de la nueva comunidad científica que está surgiendo es precisamente recuperar y discutir las ideas de Teilhard y averiguar qué pasó realmente con nuestra historia evolutiva. Solo nuevas mentes más abiertas, deseosas de saber y libres de prejuicios (figura 11) podrán avanzar en tal cometido, ya que, además, con negar la evidencia solo se conseguirá prolongar la agonía de la vieja síntesis. Sólo nuestra capacidad para formular correcta y convincentemente esta neoortogénesis de la que hablamos, podrá sustituir la cada vez más vigente (por irrebatida) visión de Teilhard de Chardin.

*******
Bibliografía
Adoutte, A., Balavoine, G, Lartillot, N, de Rosa, R. 1999. Animal evolution: the end of intermediate taxa. Trends in Genetics (15): 104108.
Albert, P. 1998. El concepto de progreso y la búsqueda de teorías generales en la evolución. En: El progreso, ¿un concepto acabado o emergente? (2021 octubre, 1995. Barcelona, España). Tusquets Editores: 193222. Barcelona.
Alonso, C. J. 1999. Tras la evolución. Panorama histórico de las teorías evolucionistas. Ediciones Eunsa. Navarra: 314 p.
Anadón, E. 1982. Sobre las dificultades para explicar la evolución. Rev. Biol. Univ. Oviedo, 1(0): 1323.
Aparicio, S. 2000. Vertebrate evolution: recent perspective from fish. Trends in Genetics (16): 5456.
Baquero, F., Blázquez, J. y Martínez, J.L. 2002. Mutación y resistencia a los antibióticos. Investigación y Ciencia (315): 7278.
Cairns, J., Overbaugh, J. & Miller, S. 1988. The origin of mutants. Nature (335): 142145.
Darwin, C. 1859. El origen de las especies. Ediciones Akal S.A. Madrid. Edición de 1994: 573 pp.
Davis, G.K., and Patel, N.H. 2000. The origin and evolution of segmentation. Trends in Genetics (15) : M68M72.
Davis, G.K. and N.H. Patel (2002). Short, long and beyond: molecular and embryological aproaches to insect segmentation. Annual Reviews of Entomology (47):669699.
De Chardin, T. 1958. El fenómeno humano. Revista de Occidente. Madrid: 334 pp.
Dobzhansky, T. 1985. La idea de especie después de Darwin. En Un siglo después de Darwin. La evolución. Alianza Editorial. Madrid: 3782.
Doménech, J.L. 1999. Evolución regresiva del Homo sapiens. Una nueva hipótesis evolutiva. Ed. J.L. Doménech. Gijón: 231 pp.
Doménech, J.L. 2004. La evolución de la consciencia a la luz de la hipótesis regresiva. Instituto de Investigación sobre la Evolución Humana A.C.
(http://www.iieh.org/doc/doc200403270003.html); último acceso: febrero/2005.
Doménech, J.L. 2005. El «origen remoto del Homo sapiens»: una teoría alternativa de la evolución humana. Congreso Internacional «La ciencia y el humanismo en el siglo XXI: perspectivas«; CDROM.
Doménech, J.L. en preparación. La «reina Alicia», una explicación para la ortogénesis evolutiva.
Doolittle, W.F. 1999. Phylogenetic Classification and the Universal Tree. Science (284): 21242128.
Gagnier, P.Y, Blieck, A.R. & Rodrigo, S.G. 1986. First Ordovician vertebrate from Sauth America. Geobios 19 (5): 629634.
Gallardo, M.H. 2000. Mamífero tetraploide. Investigación y Ciencia (289): 3839.
Goodwin, B. 1998. Forma y transformación. La lógica del cambio evolutivo. En: El progreso, ¿un concepto acabado o emergente? (2021 octubre, 1995. Barcelona, España). Tusquets Editores: 137163. Barcelona.
Gould, S. J. 1999. La vida maravillosa. Editorial Crítica, Barcelona: 357 pp.
Gould, S. J. 1994. La evolución de la vida en la Tierra. Investigación y Ciencia (219): 5461.
Grassé, P.P. 1976. Zoología. 1. Invertebrados. TorayMasson S.A. Barcelona: 938 pp.
Greenwood, P.H., Rosen, D.E., Weitzman, S.H. and Myers, G.S. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bulletin of the American Museum of Natural History. Vol. 131. Art. 4: 341455.
Herbert, A. & Rich, A. 1999. RNA processing and the evolution of eukaryotes. Nature Genetics (21): 265270.
Jacob, F., Monod, J, Calvin, M., Illya Prigogine y otros. 1986. Biología Molecular. Ediciones Orbis S.A., Barcelona: 256 pp.
JoséLeonel, Torres. 1995. En el nombre de Darwin. Consejo Nacional de Ciencia y Tecnología, Fondo de Cultura Económica de México.
Landman, E.O., 1993. Pero, ¿existe la herencia lamarckiana?. Investigación y Ciencia (202): 9696.
Lewontin, R. 1979. La adaptación. Libros de Investigación y Ciencia. En: La Evolución. Libros de Investigación y Ciencia. Editorial Labor. Barcelona: 140151. Losos, J.B., Jackman, T.R., Larson, A., Queiroz, K., RodríguezSchettino, L. 1998.
Contingency and Determinism in Replicated Adaptative Radiations of Islands Lizards. Science (279): 21152118.
Margulis, L. y Sagan, D. 1995. Microcosmos. Tusquets Editores. Barcelona: 317pp.
Margulis, L. y Sagan, D. 1999. ¿Qué es el sexo?. Tusquets Editores. Barcelona: 256pp.
Margulis, L. y Sagan, D. 2003. Captando genomas. Editorial Kairós. Barcelona: 308pp.
Mayr, E. 1979. Evolución. Libros de Investigación y Ciencia. Evolución: 212.
Mckinney, M., 1998. La escalera estadística de la evolución: el desarrollo embrionario como generador de complejidad. En: El progreso, ¿un concepto acabado o emergente? (2021 octubre, 1995. Barcelona, España). Tusquets Editores: 267306. Barcelona.
Melendez, B. 1986. Paleontología. Tomo 2. Vertebrados. Peces, Anfibios, Reptiles y Aves. Ed. Paraninfo. Madrid. 571 pp.
Mittler, J. & Lenski, R., 1990. New data on excisions of Mu from E. coli Cast Doubt on Directed Mutation Hypothesis. Nature (344).
Morell, V. 1997. Biodiversity in a Vial of Sugar Water. Science (278): 390390.
Muñoz, G.M. 2002. Origen de la vida. Generación espontánea de aminoácidos en el espacio. Investigación y Ciencia (315): 3233.
Rainey, P.B. & Travisano, M. 1997. Adaptative radiation in a heterogeneous environment. Nature (394): 6972.
Sandín, M. 2003. Sucesos excepcionales de la evolución. Instituto de Investigación sobre la Evolución Humana A.C. (http://www.iieh.org/doc/doc200311130001.html); último acceso: febrero/2005.
Sandín, M., Agudelo, G. y Alcalá, J.G. 2003. Evolución: un nuevo paradigma. Ediciones IIEH. Madrid: 256 pp.
Sheldrake, R. 1994. El renacimiento de la naturaleza. Ediciones Paidós. Barcelona: 264 pp.
Stebbins, G. L. y Ayala, F. J. 1985. La evolución del darwinismo. Investigación y Ciencia (108): 4253.
Valentine, J. W. 1979. La evolución de las plantas y los animales pluricelulares. En: La evolución. Libros de Investigación y Ciencia. Editorial Labor. Barcelona: 7083.
Valiente, E.F. 2002. Hacia un nuevo concepto de evolución». Arbor (677): 1740.
Valladares, F. 2001. Luz y evolución vegetal. Investigación y Ciencia (303): 7379.
Van Valen, L. 1973. A new evolutionary law. Evolutionary Theory (1): 130.
Wagensberg, J. 1998. El progreso, ¿un proceso acabado o emergente?. En: El progreso, ¿un concepto acabado o emergente? (2021 octubre, 1995. Barcelona, España). Tusquets Editores: 1554. Barcelona.
Whiting, M.F., Young, B. y otros. 2003. Loss and recovery of wings in stick insects. Nature (421): 264267.
INDICE- Teilhard de Chardin- EL FENOMENO HUMANO (libro completo)





Deja tu opinión