
Universo Mecánico 23 Energía y Excentricidad HD720p H 264 AAC
¿Qué es la vida? (Parte III)
CAPÍTULO 2
EL MECANISMO DE LA HERENCIA
Das Sein ist ewig; denn Gesetze Bewahren die lebendigen
Schätze, Aus welchen sich das Ah geschmückt.[1]
Goethe
2.1. La suposición del físico clásico, lejos de ser trivial, es errónea
Hemos llegado a la conclusión de que un organismo, y todos los procesos biológicos importantes que experimente, deben tener una estructura marcadamente «multi-atómica» y tienen que ser protegidos de los acontecimientos «monoatómicos» aleatorios que pudieran alcanzar una importancia excesiva. Esto, según el «físico ingenuo», es esencial para que el organismo pueda poseer leyes físicas suficientemente precisas en las que apoyarse para elaborar su maravillosamente regular y bien ordenado funcionamiento. Ahora bien, ¿cómo encajan estas conclusiones obtenidas, hablando biológicamente, a priori (es decir, desde una perspectiva puramente física) con los hechos biológicos reales?
A primera vista, uno se inclinaría a pensar que estas conclusiones son poco más que triviales. Hace treinta años, un biólogo podría haber dicho que, aun siendo oportuno que en una conferencia de divulgación se hiciera notar la importancia, en el organismo o en cualquier parte, de la Física estadística, tratábase en realidad de un lugar común familiar. Porque es evidente que tanto el cuerpo adulto de un organismo de cualquier especie superior como cada una de las células que lo componen contienen un número cósmico de átomos de todos los tipos. Y todo proceso fisiológico particular que observamos, ya sea en el interior de la célula o en la relación de esta con el ambiente, aparenta (o aparentaba al menos hace treinta años) abarcar cantidades tan enormes de átomos y procesos atómicos individuales que respetarían todas las leyes físicas y fisicoquímicas importantes, incluso teniendo en cuenta las estrictas exigencias de la Física estadística por lo que afecta a los grandes números. Exigencias que acabo de ilustrar con la regla de la √n.
Hoy en día sabemos que esta opinión habría sido errónea. Como veremos en breve, grupos increíblemente pequeños de átomos, excesivamente reducidos para atenerse a las leyes estadísticas, desempeñan de hecho un papel dominante en los ordenados y metódicos acontecimientos que tienen lugar dentro de un organismo vivo. Controlan las particularidades macroscópicas observables que el organismo adquiere en el curso de su desarrollo. Determinan importantes características de su funcionamiento, y en todo esto se manifiestan leyes biológicas muy definidas y exactas.
Debo empezar haciendo un breve sumario de la situación en la Biología, concretamente en la genética; en otras palabras, debo resumir la situación actual del conocimiento de un tema en el cual no soy experto. Esto no puedo remediarlo y pido disculpas, especialmente a los biólogos, por el carácter superficial de mi resumen. Por otra parte, permítaseme exponer las ideas básicas de forma más o menos dogmática. No puede esperarse que un pobre físico teórico haga nada parecido a una revisión competente de la evidencia experimental, que está formada por largas series, magníficamente entrelazadas, de experimentos de cruzamientos, logrados con una ingenuidad realmente sin precedentes por un lado, pero disponiendo, por el otro, de observaciones directas de la célula viva llevadas a cabo con todo el refinamiento de la microscopía moderna.
2.2. El mensaje cifrado de la herencia (cromosomas)
Permítaseme el usar el término esquema (pattern) de un organismo en el sentido en que el biólogo lo haría al decir el esquema en cuatro dimensiones, refiriéndose no solo a la estructura y funcionamiento de ese organismo en el estado adulto, o en cualquier otra fase particular, sino al conjunto de su desarrollo ontogénico, desde el huevo fertilizado al estado de madurez, cuando el organismo empieza a reproducirse.
Ahora bien, todo este esquema en cuatro dimensiones esta determinado en la estructura de esa célula única que es el huevo fertilizado. Más aún, sabemos que está determinado esencialmente por solo una parte de esta célula, su núcleo. Este núcleo, en el «estado de reposo» normal, aparece, por regla general, como una masa de cromatina.[2] Pero, en los procesos de división celular, de vital importancia (denominados mitosis y meiosis, como veremos más adelante), se observa que está constituido por un conjunto de partículas, en general con forma de fibras o bastones, llamados cromosomas, que pueden ser 8, 12, etc.; en el hombre son 46. En realidad, debería haber escrito estos ejemplos como 2×4, 2×6…, 2×23…, y tendría que haber hablado de dos conjuntos o dotaciones, para utilizar la expresión tal como lo hacen los biólogos. Ya que, aunque los cromosomas individuales son a veces claramente diferenciables por tamaño y forma, existen dos conjuntos que son prácticamente idénticos. Como veremos en seguida, un conjunto proviene de la madre (ovulo) y el otro del padre (espermatozoide fertilizador). Son estos cromosomas, o probablemente solo una fibra axial de lo que vemos bajo el microscopio como cromosoma, los que contienen en alguna forma de clave o texto cifrado el esquema completo de todo el desarrollo futuro del individuo y de su funcionamiento en estado maduro. Cada dotación completa de cromosomas contiene toda la clave; de este modo, por regla general son dos copias de esta las que encontramos en el huevo fertilizado, el cual constituye el primer estadio del individuo futuro.
Al decir que la estructura de las fibras de los cromosomas son un texto cifrado queremos significar que la inteligencia absoluta, imaginada por Laplace, para la que cualquier relación causal sería evidente, podría averiguar, partiendo de su estructura, si de un huevo, bajo determinadas condiciones, se desarrollaría un gallo negro o una gallina moteada, una mosca o una planta de maíz, un rododendro, un escarabajo, un ratón, o una mujer. A esto podríamos añadir que el aspecto de las distintas células-huevo es muy similar; y que, aun cuando no lo sea, como en el caso de los gigantescos, en comparación, huevos de reptiles y aves, la diferencia no radica en las estructuras importantes, sino en la cantidad de material nutritivo que se añade en estos casos por razones obvias.
Pero el término clave, o texto cifrado, es demasiado limitado. Las estructuras cromosómicas son al mismo tiempo los instrumentos que realizan el desarrollo que ellos mismos pronostican. Representan tanto el texto legal como el poder ejecutivo; para usar otra comparación, son a la vez los planos del arquitecto y la mano de obra del constructor.
2.3. Crecimiento del cuerpo por división celular (mitosis)
¿Cuál es el comportamiento de los cromosomas a lo largo de la ontogenia?[3]
El crecimiento de un organismo se lleva a cabo por divisiones celulares consecutivas. Cada división celular de este tipo se denomina mitosis. En la vida de una célula, este es un acontecimiento menos frecuente de lo que cabría esperar, considerando la enorme cantidad de células que componen nuestro cuerpo. Al principio, el crecimiento es rápido. La célula-huevo se divide en dos «células hijas», que darán lugar, en la siguiente etapa, a una generación de cuatro células, después 8, 16, 32, 64, etc. La frecuencia de división no se mantiene constante en todas las partes del individuo en crecimiento, y así se altera la regularidad de estos números. Pero de su rápido aumento deducimos, por un cálculo fácil, que por término medio bastan solo 50 o 60 divisiones sucesivas para producir el número de células[4] que encontramos en un hombre adulto, o incluso un número diez veces mayor, considerando también la renovación de células a lo largo de su vida. Así pues, una célula de mi cuerpo es, como media, tan solo el quincuagésimo o sexagésimo descendiente de la célula huevo que yo fui.
2.4. En la mitosis se duplica cada uno de los cromosomas
¿Cómo se comportan los cromosomas en la mitosis? Se duplican; las dos dotaciones, las dos copias de la clave, se duplican. El proceso ha sido minuciosamente estudiado bajo el microscopio y es de gran interés, pero excesivamente complicado como para exponerlo aquí con detalle. El principal hecho es que cada una de las «células hijas» recibe una dotación completa de cromosomas exactamente igual a la de la «célula madre». De este modo, todas las células del cuerpo son idénticas por lo que se refiere a su tesoro cromosómico[5].
Por poco que sepamos de este sistema, es evidente que debe de resultar muy importante para el funcionamiento del organismo, ya que todas y cada una de las células, incluso la menos importante, debe poseer una copia completa (doble) del texto cifrado. El general Montgomery, en su campaña africana, consideraba fundamental que cada uno de los soldados de su ejercito estuviera meticulosamente informado de sus planes. Si esto fue así (y probablemente lo fuera, considerando el nivel de inteligencia y fiabilidad de sus tropas), encontramos una analogía con el caso que nos ocupa, en el que el hecho se cumple literalmente. Lo más sorprendente es que la dotación cromosómica sea doble, manteniéndose así a través de las divisiones mitóticas. Que esto es la característica más destacada de un mecanismo genético queda revelado, de la manera más sorprendente, por la única excepción a la regla, como veremos a continuación.
2.5. División reductora (meiosis) y fertilización (singamia)
Muy poco después de empezar el desarrollo del individuo, un grupo de células queda reservado para producir, en un estadio posterior, lo que llamamos gametos, los espermatozoides u óvulos, según sea el caso, que son necesarios para la reproducción de un individuo en la madurez. Al decir reservado, nos referimos al hecho de que estas células no son utilizadas entretanto para otras finalidades y sufren muchas menos divisiones mitóticas que las demás células. La división excepcional o reductora (denominada meiosis) es aquella por la cual, cuando el organismo alcanza la madurez, se producen los gametos a partir de las células reservadas; por lo general, solo poco tiempo antes de que tenga lugar la singamia. Por la meiosis, la doble dotación de cromosomas de la célula madre se separa y cada una va a parar a una célula hija, los gametos, que se forman siempre de cuatro en cuatro. En la meiosis se produce una duplicación del número de cromosomas, pero estos se reparten entre cuatro células, por lo que cada una de las cuatro recibe solo la mitad, es decir, solo un ejemplar completo de la clave en lugar de dos. Por ejemplo, en el hombre, los gametos tienen 23 cromosomas, y no 2×23 = 46.
Las células con solo una dotación cromosómica se llaman haploides (del griego απλους, único). Por lo tanto los gametos son haploides, y las células comunes del cuerpo diploides (del griego διπλους, doble). A veces, se encuentran individuos con tres, cuatro… o muchas dotaciones cromosómicas en todas las células de su cuerpo; se les denomina triploides, tetraploides… o poliploides.
En el acto de la singamia, el gameto masculino (espermatozoide) y el gameto femenino (óvulo), ambas células haploides, se reúnen para formar la célula huevo fertilizada, que, en consecuencia, es diploide. Una de sus dotaciones cromosómicas proviene del padre y la otra de la madre.
2.6. Individuos haploides
Se nos presenta otro punto que requiere una rectificación. Aunque para nuestro objetivo no es indispensable, resulta muy interesante, ya que permite ver que en una sola dotación cromosómica se encuentra realmente de forma completa el texto cifrado del esquema del individuo.
Hay casos en los que la meiosis no es seguida poco después de la fertilización, y la célula haploide sufre numerosas divisiones mitóticas, lo cual da lugar a un individuo haploide completo. Así ocurre, por ejemplo, en la abeja macho, el zángano, que es producido partenogenéticamente, es decir, a partir de óvulos no fertilizados, y por lo tanto haploides, de la reina. ¡El zángano no tiene padre! Todas las células de su cuerpo son haploides. Podría decirse que es un espermatozoide exageradamente grande; y, de hecho, como todo el mundo sabe, esa es su única misión en la vida. Sin embargo, tal vez sea este un punto de vista algo cómico, ya que el caso no es único. En algunos grupos de plantas, la célula haploide, producida por meiosis y denominada espora, cae al suelo y da lugar, como si fuera una semilla, a una planta completa, haploide, de tamaño comparable al de la diploide. La figura 1 es el esquema de un musgo bien conocido en nuestros bosques. La parte inferior, foliada, es la planta haploide, denominada gametofito, ya que en su parte superior desarrolla órganos sexuales y gametos, que por mutua fertilización producen en forma habitual la planta diploide, que es el tallo desnudo con una capsula en el extremo. Esta parte se denomina esporofito, porque produce, por medio de la meiosis, las esporas que la capsula contiene. Cuando la capsula se abre, las esporas caen al suelo y dan lugar al tallo foliado. Y así sucesivamente. A esa sucesión de acontecimientos se la ha llamado, de forma muy apropiada, alternación de generaciones. Si quisiéramos, también podríamos hablar en estos términos del caso más común del hombre y los animales. Pero entonces el «gametofito» sería una breve generación unicelular, espermatozoide u óvulo, según el sexo.
Nuestro cuerpo correspondería al esporofito, nuestras «esporas» las células reservadas de las que, por meiosis, se origina la generación unicelular de gametos.

2.7. La extraordinaria importancia de la división reductora
El hecho importante y realmente decisivo en la reproducción del individuo no es la fertilización, sino la meiosis. Una colección de cromosomas proviene del padre, otra de la madre. Ni la casualidad ni el destino pueden alterar ese hecho. Cada hombre[6] debe a su madre exactamente la mitad de su herencia genética y a su padre la otra mitad. El hecho de que a menudo parezca que domine una u otra parte se debe a otras razones, como veremos más adelante. (El sexo es, de hecho, el aspecto más simple y constante de tal predominio.)
Pero, cuando seguimos la pista de nuestra herencia hasta nuestros abuelos, la situación es distinta. Fijémonos, por ejemplo, en mi colección paterna de cromosomas, en particular en uno de ellos, digamos el número 5. Éste es una réplica fiel del número 5 que mi padre recibió de su padre o del número 5 que recibió de su madre. El resultado fue decidido con una probabilidad de un 50% en la meiosis que tuvo lugar en el cuerpo de mi padre en noviembre de 1886, la cual produjo el espermatozoide que unos días después se haría efectivo al engendrarme a mí. Podríamos repetir la misma historia para el cromosoma 1, 2, 3, …, 23 de mi colección paterna, y mutatis mutandis para cada uno de mis cromosomas maternos. Por otra parte, cada una de las 46 soluciones es independiente de las otras. Es decir, incluso sabiendo que mi cromosoma paterno número 5 proviene de mi abuelo Josef Schrödinger, el número 7 sigue teniendo la misma probabilidad de venir o bien de él, o bien de su mujer, Marie Bogner.
2.8. Entrecruzamiento. Localización de los caracteres
Sin embargo, el azar tiene aún más recursos para mezclar la herencia recibida por el nieto de sus abuelos que los que hemos visto hasta ahora. Hasta aquí hemos supuesto tácitamente, o incluso dicho, que un cromosoma determinado proviene como un todo del abuelo o de la abuela: es decir, que los cromosomas se transmiten indivisos, de una sola pieza. En realidad, no es así, o por lo menos no siempre es así. Antes de ser separados por la división reductora, por ejemplo la del cuerpo del padre, cada pareja de cromosomas «homólogos» entra en estrecho contacto, durante el cual intercambia a veces trozos enteros, como podemos ver en la figura 2. Por este proceso, denominado entrecruzamiento (crossing-over), dos propiedades que estaban situadas en dos partes diferentes de un mismo cromosoma estarán separadas en el nieto, que será como el abuelo en una parte y como la abuela en otra. La existencia del entrecruzamiento, que no es muy frecuente pero tampoco excepcional, nos ha suministrado inapreciable información acerca de la localización de los caracteres en los cromosomas. Para hacer una exposición completa, deberíamos profundizar en conceptos que no introduciremos hasta el próximo capítulo (por ejemplo, heterozigosis, dominancia, etc.); pero, como esto nos llevaría más allá de lo que se pretende en este pequeño libro, permítaseme indicar solo el aspecto principal ahora mismo.
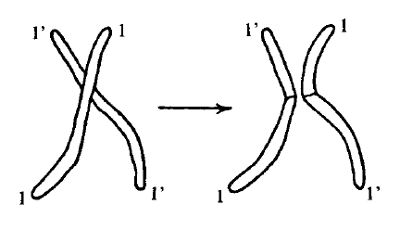
Si no hubiera entrecruzamiento, dos caracteres de los que fuera responsable un mismo cromosoma se transmitirían siempre juntos y ningún descendiente recibiría uno sin el otro. Por otra parte, dos caracteres debidos a cromosomas distintos, o bien tendrían una probabilidad de un 50% de estar separados o bien estarían siempre separados (esto último en el caso de estar situados en cromosomas homólogos de un mismo antepasado, los cuales no podrían reunirse nunca).
El entrecruzamiento altera reglas y probabilidades. En consecuencia, registrando cuidadosamente el porcentaje de la composición de la prole en experimentos con abundantes cruces experimentales, y convenientemente preparados, podremos conocer la probabilidad del fenómeno. Analizando los resultados estadísticos, se acepta la sugestiva hipótesis de trabajo según la cual el ligamento (linkage) entre dos caracteres situados en el mismo cromosoma se rompe, por entrecruzamiento, tanto menos frecuentemente cuanto más cerca están los caracteres entre sí. Esto se debe a que el punto de intercambio tiene una menor probabilidad de encontrarse entre ellos, mientras que los caracteres que estén localizados cerca de los extremos opuestos del cromosoma serán separados por todos los entrecruzamientos que se produzcan. (Con la recombinación de caracteres, situados en cromosomas homólogos del mismo antecesor, ocurre algo parecido.) De esta forma, se puede esperar que, a partir de las «estadísticas de ligamiento», se obtenga una especie de «mapa de caracteres» dentro de cada cromosoma.
Estas suposiciones se han confirmado plenamente. En los casos en que se han hecho ensayos cuidadosos (principal pero no exclusivamente, en la mosca del vinagre, drosophila), los caracteres analizados se reparten en grupos independientes, sin que haya ligamiento entre un grupo y otro, correspondiendo cada grupo a los diferentes cromosomas (cuatro en drosophila). Dentro de cada grupo, puede elaborarse un mapa lineal de los caracteres dando una valoración cuantitativa del grado de ligamiento entre dos cualesquiera de ese grupo, de modo que no haya duda acerca de los puntos en los que se localizan realmente, a lo largo de una línea, tal como sugiere la forma de bastón del cromosoma.
Por supuesto, el esquema del mecanismo de la herencia que hemos esbozado hasta aquí es bastante vacío y descolorido, incluso ingenuo. Por ejemplo, no hemos dicho que entendemos exactamente por «carácter». No parece posible ni adecuado diseccionar en «caracteres» la estructura de un organismo que es esencialmente una unidad, «un todo». Ahora bien, lo que decimos en cada caso particular es que un par de antepasados se diferenciaba en algún aspecto bien definido (por ejemplo, uno tenía ojos azules y el otro castaños) y que su descendencia se parece en este aspecto a uno o a otro. Lo que situamos en el cromosoma es la localización de esta diferencia. (En lenguaje técnico, lo llamaríamos un locus, o, si pensamos en la hipotética estructura material que lo constituye, un gen.) Desde mi punto de vista, la diferencia en el carácter es el concepto fundamental, más que el carácter en sí, a despecho de la aparente contradicción lingüística y lógica de tal afirmación. Las diferencias entre los caracteres son en realidad discretas, tal como se verá en el próximo capítulo cuando hablemos de las mutaciones; espero que, entonces, el árido esquema presentado hasta ahora, adquirirá más vida y color.
2.9. Tamaño máximo de un gen
Acabamos de presentar el gen como el hipotético transportador material de una determinada característica hereditaria. Ahora debemos hacer notar dos puntos que resultaran de gran importancia para nuestra investigación. El primero es el tamaño (o más exactamente, el tamaño máximo) de dicho transportador. En otras palabras, ¿hasta qué diminuto volumen podemos detectar su localización? El segundo punto será la permanencia de un gen, que inferiremos de la durabilidad de la estructura hereditaria.
Por lo que hace al tamaño, se dispone de dos estimaciones totalmente independientes, una basada en pruebas genéticas (cruces experimentales) y la otra en pruebas citológicas (inspección microscópica directa). La primera es, en principio, bastante sencilla. Comenzamos situando en el cromosoma, por el sistema descrito antes, un número considerable de caracteres diferentes (por ejemplo, los de la mosca drosophila). Para obtener la estimación que se busca, en un cromosoma en concreto, bastara con dividir la longitud medida de ese cromosoma por el número de características y multiplicarla por la sección transversal. Evidentemente, consideraremos solo como diferencias aquellas características que alguna vez son separadas por entrecruzamiento, de forma que no puedan ser debidas a la misma estructura (microscópica o molecular). Está claro, por otra parte, que nuestra estimación solo podrá dar un tamaño máximo, ya que el número de características individualizadas por el análisis genético aumenta continuamente a medida que avanza la investigación.
La otra estimación, a pesar de estar basada en observaciones microscópicas, es mucho menos directa. Algunas células de drosophila (concretamente las de las glándulas salivares de las larvas) están, por algún motivo, enormemente aumentadas, y lo mismo ocurre con sus cromosomas. En ellos se distingue un apretado esquema de bandas oscuras transversales. C. D. Darlington observó que el número de estas bandas (2000 en el caso que el estudio) es, aunque considerablemente mayor, de aproximadamente el mismo orden de magnitud que el número de genes localizados en aquel cromosoma por los cruces experimentales. Lo cual le hace considerar las bandas como indicadores de los propios genes (o de la separación entre ellos). Dividiendo la longitud del cromosoma de una célula de tamaño normal por ese número (2000) se encuentra que el volumen de un gen es el de un cubo de 300 Å de arista. Teniendo en cuenta el carácter aproximado de las estimaciones, es el mismo resultado que el obtenido por el primer método.
2.10. Números pequeños
Más adelante discutiremos a fondo la relación de lo visto hasta aquí con la Física estadística, o quizá, debería decir, la relación de estos hechos con la aplicación de la Física estadística a la célula viva. Pero permítaseme hacer notar el hecho de que 300 Å son solo unas 100 o 150 distancias atómicas en un líquido o en un sólido, de forma que un gen con toda seguridad no contiene más que un millón o unos pocos millones de átomos. Esta cifra es demasiado pequeña (desde el punto de vista de la √n) para suponer un comportamiento regular y ordenado según la Física estadística (y esto equivale a decir que según toda la Física). Es excesivamente pequeño, incluso si todos estos átomos desempeñaran funciones idénticas, como ocurre en un gas o en una gota de líquido. Y evidentemente el gen es algo más que una homogénea gota de líquido. Probablemente se trata de una gran molécula de proteína, en la que cada átomo, cada radical, cada anillo heterocíclico, tiene un papel individual y más o menos distinto del que tiene cualquiera de los otros átomos, radicales o anillos. Ésta es, al menos, la opinión de genetistas de vanguardia como Haldane y Darlington, y pronto haremos referencia a experimentos genéticos que lo demuestran[7].
2.11. Permanencia
Vayamos ahora al segundo punto importante: ¿cuál es el grado de permanencia que encontramos en los caracteres hereditarios y que conclusiones debemos sacar sobre las estructuras materiales que los transportan?
La respuesta podemos darla sin ninguna investigación especial. El mero hecho de que hablemos de propiedades hereditarias indica que reconocemos esta permanencia como prácticamente absoluta. No debemos olvidar que lo que se transmite de padres a hijos no es esta o esa peculiaridad concreta, nariz aguileña, dedos cortos, tendencia al reumatismo, hemofilia o polidactilia. Éstas son características que podemos seleccionar para estudiar las leyes de la herencia. Pero, en realidad, es la estructura completa (en cuatro dimensiones) del fenotipo, la naturaleza visible y manifiesta del individuo, lo que se reproduce sin cambios apreciables a través de las generaciones, permanente a lo largo de los siglos (aunque no a lo largo de decenas de miles de años) y aportada en cada transmisión por la estructura material de los núcleos de las dos células que se unen para formar el huevo fertilizado. Eso es una maravilla, solo superada por otra, la cual, si bien íntimamente relacionada con ésta, se encuentra a otro nivel. Me refiero al hecho de que nosotros, que estamos en todo nuestro ser basados en unas relaciones prodigiosas de este tipo, tenemos además el poder de adquirir un considerable conocimiento de ello. Creo posible que este conocimiento llegue muy cerca de una comprensión completa de la primera maravilla. La segunda probablemente se encuentre fuera del alcance de la mente humana.
♦♦♦♦♦♦♦
[1] El ser es eterno; existen leyes para conservar los tesoros de la vida, de las cuales el Universo extrae su belleza.
[6] Por supuesto, también cada mujer. Para evitar disgresiones he excluido de este resumen el interesante tema de la determinación del sexo y de los caracteres ligados a este (por ejemplo, la ceguera para los colores).
Deja tu opinión